§ 16. ПЕРЕНОС ГЕНЕТИЧЕСКОЙ ИНФОРМАЦИИ В КЛЕТКЕ: ТРАНСляцИЯ
Синтез белка (трансляция) в клетке представляет собой, пожалуй, самый сложный биосинтетический процесс. В нем участвует очень большое число белков, иРНК, тРНК, рРНК в составе рибосом и другие молекулы. При его протекании затрачивается большое количество энергии. Различают несколько стадий биосинтеза белка: активация аминокислот, инициация, элонгация и терминация.
Соответствие между полинуклеотидной и полипептидной последовательностями
Синтез белка отличается от других матричных процессов (репликации, транскрипции) тем, что между матрицей (иРНК) и продуктом (белком) нет комплементарного соответствия. Для расшифровки нуклеотидной последовательности необходим генетический код. Он устанавливает соответствие между нуклеотидной последовательностью иРНК и синтезируемой на ней полипептидной цепью. Единицей генетического кода является кодон. Кодон представляет собой последовательность, состоящую из трех нуклеотидов, т.е. триплет. Всего существует 64 кодона. Из них 61 кодон используется для кодирования аминокислот. Три же кодона не кодируют ни одну из аминокислот и служат сигналом для остановки синтеза полипептидной цепи. Это так называемые терминирующие, или нонсенс-кодоны. Каждому кодону (из 61) соответствует строго определенная аминокислота, например, триплету УУУ соответствует аминокислота фенилаланин (таблица 6), т.е. код однозначен. Следовательно, зная последовательность иРНК, можно определить аминокислотную последовательность закодированного в ней полипептида:
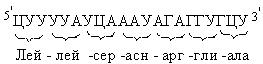
Трансляция иРНК (считывание информации) начинается с инициирующего триплета – АУГ, и далее расшифровывается каждый последующий триплет в направлении от 5’-конца молекулы иРНК к 3’-концу, заканчивается синтез полипептида на одном из трех терминирующих кодонов (рис. 52). Синтез же полипептидной цепи начинается с N-конца.
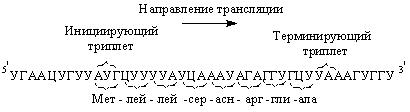
Рис. 52. Кодирующая последовательность начинается с инициирующего триплета и заканчивается терминирующим
Как ранее отмечалось, существуют 20 стандартных аминокислот. Этим 20 аминокислотам соответствует 61 кодон. Таким образом, почти каждой стандартной аминокислоте соответствует несколько кодонов, т.е. одна аминокислота может быть закодирована несколькими кодонами. Из этого следует, что нельзя однозначно перевести аминокислотную последовательность данного белка в нуклеотидную последовательность иРНК.
Таблица. 6
Генетический код
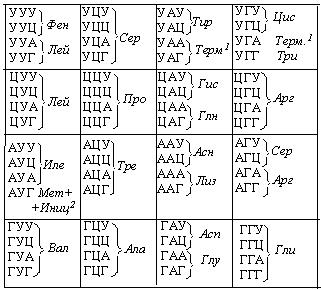
Примечание: Терм.1 – терминирующий кодон
Иниц.2 – инициирующий кодон
Информационная РНК
Информационные РНК (их еще называют матричные РНК (мРНК)) служат матрицами для биосинтеза полипептидных цепей. Они содержат линейную последовательность кодонов, которые и определяют первичную структура белка. иРНК – это одноцепочечные молекулы. Одна молекула иРНК может кодировать одну или несколько полипептидных цепей. Если иРНК несет информацию об одной полипептидной цепи, то ее называют моноцистронной, если о двух или более – полицистронной. иРНК прокариот бывают часто полицистронными, иРНК эукариот являются моноцистронными. На 3’- и 5’- концах иРНК содержат некодирующие последовательности. Полицистронные иРНК также могут содержать нетранслируемые межгенные области, которые разделяют участки, кодирующие полипептидные цепи. иРНК эукариот на 5’-конце имеет кэп, а на 3’- конце – полиА. На рис. 53 представлены схемы строения иРНК прокариот и эукариот.
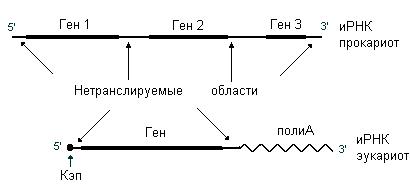
Рис. 53. Информационные РНК
Транспортные РНК
тРНК трансформируют генетическую информацию, закодированную в иРНК, в информацию о первичной структуре белка.
тРНК – это небольшие молекулы, состоящие из 73 – 93 нуклеотидов, что соответствует относительной молекулярной массе 24000 – 31000. Каждой аминокислоте соответствует одна или более тРНК. На рис. 54 показано строение тРНК. Молекула тРНК имеет вид клеверного листа. Между азотистыми основаниями в ее молекуле образуются водородные связи. На 3’-конце всех тРНК находится тринуклеотидная последовательность Ц-Ц-А. В тРНК выделяют акцепторную и антикодоновую ветви. К акцепторной ветви присоединяется аминокислота. А антикодоновая ветвь содержит антикодон, - триплет нуклеотидов, который комплементарен соответствующему кодону иРНК. Более подробно о назначении акцепторной ветви и антикодона поговорим чуть позже.
Интересно знать! Обнаружены тРНК, которые обусловливают нестандартное считывание кодовой таблицы, причем антикодоны этих тРНК некомплементарны считываемым кодонам. Обнаружены и альтернативы в чтении кода. Так терминирующий кодон УГА у разных объектов кодирует необычную аминокислоту – селено-цистеин, но только при условии, что этот кодон оказывается в определенной точке гена.
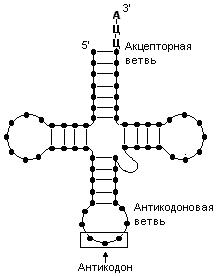
Рис. 54. тРНК
Рибосомы
Рибосомы – это субклеточные структуры, являющиеся местом синтеза белка. Рибосомы состоят из двух субъединиц – большой и малой. В состав рибосом входят белки и рРНК (рис. 55). В прокариотических рибосомах присутствуют три вида рРНК, в эукариотических – 4. рРНК играют важную роль в структуре и биосинтетической функции рибосом.

Рис. 55. Рибосомы
Активация аминокислот
На этой стадии каждая из 20 аминокислот присоединяется к определенной тРНК. При этом используется энергия АТФ. Эти реакции катализируются 20 различными ферментами, называемыми аминоацил-тРНК-синтетазами. Каждая аминоацил-тРНК-синтетаза способна узнавать только одну определенную аминокислоту и соответствующую ей тРНК. Они присоединяют аминокислотный остаток к 2’- или 3’-гидроксильной группе 3’-концевого нуклеотида. Суммарная реакция активации аминокислоты выглядит так:
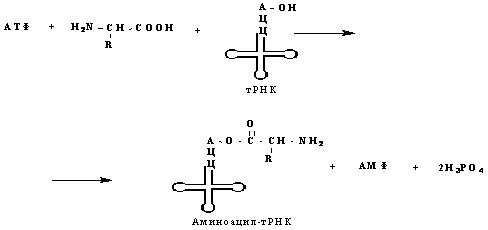
Инициация белкового синтеза
Процессы трансляции эукариотической иРНК и прокариотической иРНК в общих чертах сходны. Инициация начинается с присоединения к малой субъединице рибосомы иРНК и первой аминоацил-тРНК (аа-тРНК), антикодон которой комплементарен инициирующему кодону АУГ (рис. 56). После связывания антикодона тРНК с инициирующим кодоном происходит присоединение большой субъединицы рибосомы. Образовался инициирующий комплекс, в котором инициирующая аа-тРНК находится в Р (пептидильном)-центре, а А (аминоацильный) – центр свободен. Инициирующей аа-тРНК у эукариот является метионил-тРНК, у прокариот – формилметионил-тРНК, образующийся при модификации метионил-тРНК. Для осуществления инициации трансляции необходима энергия. Ее поставляет ГТФ. Поставляемая энергия высвобождается при гидролизе ГТФ до ГДФ и фосфата.
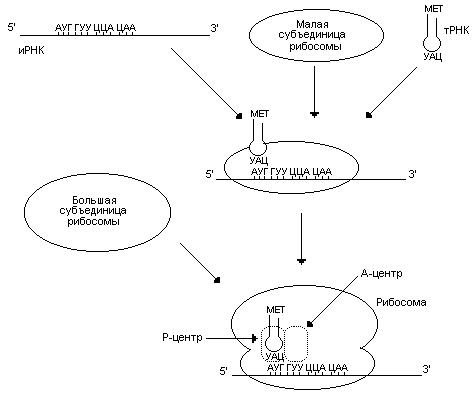
Рис. 56. Инициация трансляции. Инициирующей аа-тРНК у эукариот является метионил-тРНК
Элонгация белкового синтеза
В процессе элонгации (рис. 57) происходит наращивание полипептидной цепи. При этом присоединение каждого аминокислотного остатка происходит в три стадии. Этот цикл продолжается до тех пор, пока рибосома не достигнет терминирующего кодона. На первой стадии в А-центре происходит связывание следующей аа-тРНК, антикодон которой комплементарен следующему за инициирующим кодону. Между кодоном и антикодоном возникает комплементарное взаимодействие. Далее происходит образование пептидной связи за счет пептидилтрансферазной реакции, в результате которой метионин (у прокариот формилметионин) переносится на a-аминогруппу аа-тРНК, находящуюся в А-центре. В результате образуется дипетидил-тРНК. Катализирует этот процесс рРНК большой субъединицы рибосомы. После транспептидазной реакции рибосома перемещается на один кодон в направлении от 5’- к 3’-концу мРНК. Дипептидил-тРНК из А-центра перемещается в Р-центр, А-центр освобождается, здесь оказывается следующий кодон. Инициирующая тРНК покидает рибосому. Рибосома готова к новому циклу элонгации. Далее присоединяется третий аминокислотный остаток, потом – четвертый и т.д. По мере движения рибосомы вдоль иРНК аминокислотные остатки один за другим добавляются к растущей полипептидной цепи. В процессе трансляции затрачивается энергия, освобождающаяся при гидролизе ГТФ до ГДФ и фосфата.
Терминация
Терминация белового синтеза наступает, как только в А-центре окажется один из терминирующих кодонов: УАГ, УГА, УАА. В этом процессе участвуют специфические белки – факторы терминации. В результате терминации происходит гидролитическое отщепление полипептида от тРНК, тРНК отделяется от рибосомы, рибосома диссоциируют на субъединицы. Поставщиком энергии для терминации синтеза белка так же, как и для инициации и элонгации, является ГТФ.
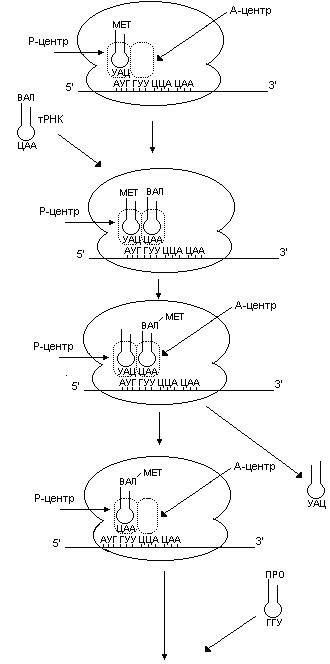
Рис. 57. Элонгация биосинтеза белка