§ 14. СТРУКТУРА НУКЛЕИНОВЫХ КИСЛОТ
Первичная структура нуклеиновых кислот
Первичная структура нуклеиновых кислот представляет собой порядок чередования нуклеотидов в полинуклеотидной цепи (рис. 40). Нуклеотиды в молекулах ДНК и РНК связаны друг с другом фосфодиэфирными мостиками между 3’- и 5’- углеродными атомами остатков пентоз.
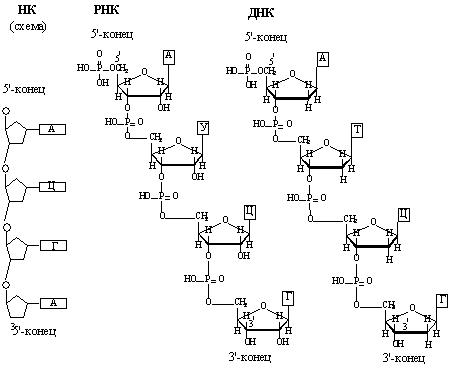
Рис. 40. Первичная структура нуклеиновых кислот
Цепи ДНК и РНК обладают полярностью, каждая цепь имеет5’и -3’-концы (рис.40). Последовательность нуклеотидов в одиночной цепи ДНК и РНК записывается в направлении от 5’-конца к 3’-концу. Нумерация нуклеотидов также начинается с 5’- конца.
Вторичная структура ДНК
Вторичная структура (двойная спираль) была предложена американским генетиком Д.Уотсоном и английским физиком Ф.Криком в1953 г. Это открытие произвело переворот в исследовании нуклеиновых кислот. Предпосылкой для постулирования структуры двойной спирали послужили исследования других ученых. Э.Чаргафф и более поздние исследователи, изучая нуклеотидный состав ДНК различных видов организмов, сделали следующие выводы:
1. нуклеотидный состав ДНК разных тканей одного и того же вида одинаков;
2. нуклеотидный состав ДНК у разных видов различен;
3. нуклеотидный состав не зависит от возраста и питания;
4. в составе ДНК число остатков аденина всегда равно числу остатков тимина, а число остатков гуанина равно числу остатков цитозина. Из этого следует, что сумма пуриновых оснований равна сумме пиримидиновых – А+Г=Т+С.
Тогда же в 50-е гг. ХХ в., Р.Франклин и М.Уилкинс опубликовали рентгенограмму, полученную при рентгеноструктурном анализе нитей ДНК. Метод рентгеноструктурного анализа широко используется при исследовании пространственной структуры молекул. Предложенная Уотсоном и Криком модель пространственной организации ДНК объяснила результаты исследований выше названых ученых.
Согласно этой модели, ДНК состоит из двух цепей, закрученных в правую двойную спираль (рис. 41б.). При этом цепи располагаются антипараллельно (рис. 41а.), т.е. они ориентированы во взаимно противоположных направлениях. Плоские молекулы азотистых оснований расположены перпендикулярно оси двойной спирали. На внешней стороне двойной спирали расположены остатки дезоксирибозы и фосфорной кислоты. Цепи ДНК связаны друг с другом водородными связями, которые образуются между гуанином одной цепи и цитозином другой цепи, а также между тимином и аденином, расположенными в разных цепях. При этом между тимином и аденином образуются две водородные связи, а между гуанином и цитозином – три водородные связи (рис. 42.). Способность гуанина взаимодействовать в молекуле ДНК только с цитозином, а аденина – только с тимином называют комплементарностью, а основания гуанин и цитозин, аденин и тимин – комплементарными. Согласно принципу комплементарности, последовательность одной цепи будет определять последовательность другой цепи. Всегда против аденина будет находится тимин, а против гуанина – цитозин. Таким образом, цепи ДНК в двойной спирали будут комплементарны друг другу. Помимо водородных связей, возникающих при взаимодействии комплементарных оснований друг с другом, второй, не менее значительной, силой, стабилизирующей двойную спираль, является стэкинг – взаимодействие находящихся в стопке оснований. Основания расположены друг над другом и сближены своими плоскостями. В результате между ними возникают гидрофобные взаимодействия, а также дипольные взаимодействия p–связей.
Цепи ДНК (рис. 41б) при закручивании в двойную спираль образуют большую и малую борозды, ширина большой борозды – 2,2 нм, малой – 1,2 нм. На один виток спирали приходится 10 нуклеотидных остатков. Полный виток спирали имеет длину 3,4 нм. Диаметр двойной спирали 1,8 нм.
Параметры двойной спирали в зависимости от условий и состава ДНК могут несколько отличатся от той модели, которую предложили Уотсон и Крик. В настоящее время описаны и другие модели ДНК. Тем не менее во всех предложенных моделях сохраняется принцип комплементарности, и цепи ДНК закручены в двойную спираль.
Интересно знать! Цепи ДНК двух различных видов могут образовывать гибриды. Если ДНК двух различных видов смешать и нагреть (при этом произойдет расхождение цепей - плавление ДНК), а затем раствор охладить и выдержать в течение определенного времени, то некоторая часть ДНК будет образовывать гибридную двойную спираль, состоящую из цепей ДНК различных видов. Чем ближе родство двух видов, тем больше будет образовываться гибридных молекул. Данный подход используется для выяснения филогенетического родства видов.
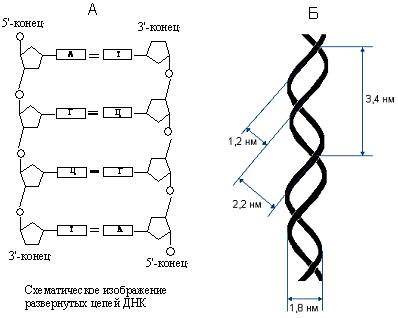
Рис. 41. Параметры ДНК.
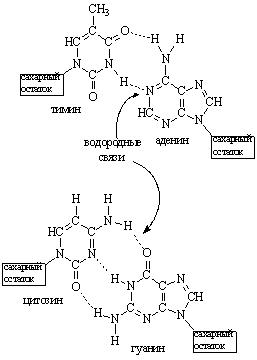
Рис. 42. Образование водородных связей между аденином и тимином, гуанином и цитозином в молекуле ДНК
Пространственная организация РНК
Природные РНК можно разделить на 2 группы: одноцепочечные и двухцепочечные РНК. Двухцепочечные РНК состоят из двух нитей РНК, комплементарных друг другу. Этот тип РНК встречается в составе некоторых вирусов. По своей организации двухцепочные РНК сходны с ДНК. Они закручены в двойную правую спираль, цепи РНК в них антипаралельны, между комплементарными основаниями образованы водородные связи, углеводнофосфатный скелет расположен снаружи спирали.
Большинство же природных РНК являются одноцепочечными. Несмотря на это, в своей структуре они могут иметь фрагменты двойной спирали, чередующиеся с линейными одноцепочечными участками РНК. Фрагменты двойной спирали образованы комплементарными участками РНК, расположенными в пределах одной цепи (рис. 43.). В некоторых случаях доля двухспиральных участков в РНК может достигать 75 – 90 %.
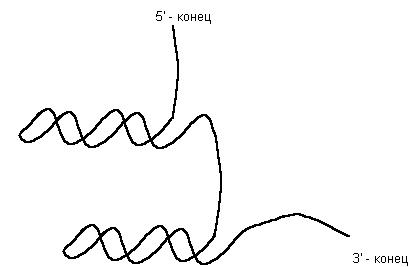
Рис. 43. Двухцепочечные шпильки в молекуле РНК