§ 15. ПЕРЕНОС ГЕНЕТИЧЕСКОЙ ИНФОРМАЦИИ В КЛЕТКЕ:
РЕПЛИКАЦИЯ, ТРАНСКРИПЦИЯ
В процессе размножения клеток генетическая информация передается от одного поколения клеток другому. При этом все клетки получают одинаковую информацию. Это, возможно, вследствие того, что перед делением клетки осуществляется репликация (удвоение) ДНК, в результате образуются две идентичные молекулы ДНК, которые и передаются потомкам. В структуре ДНК заложена способность этой молекулы к копированию.
Закодированная в ДНК генетическая информация реализуется в результате экспрессии генов. Экспрессия генов включает транскрипцию (копирование информации с ДНК на синтезируемую РНК) и последующую трансляцию (синтез на матрице РНК соответствующего белка).
Возможен поток информации и в направлении от РНК к ДНК, этот процесс носит название обратная транскрипция. В то же время информация не передается от белков нуклеиновым кислотам. Однако следует отметить, что белки играют важную роль в осуществлении процессов передачи информации, как между нуклеиновыми кислотами, так и от нуклеиновых кислот к белкам.
Описанные информационные взаимоотношения между ДНК, РНК и белками могут быть представлены в виде схемы:
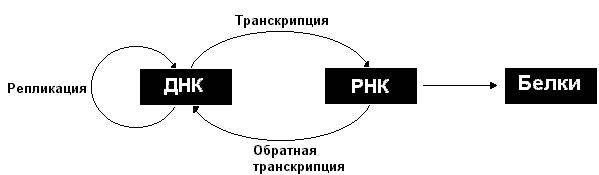
Рассмотрим более подробно процессы репликации, транскрипции и трансляции.
Интересно знать! В 1982 году Р.Д.Пальмитер с коллегами опубликовали фотографию, на которой рядом сидели две мыши. Одна из них была трансгенной, в ее ДНК встроили ген гормона роста крысы, другая была обычной мышкой. Трансгенная мышь в сравнении с обычной имела в два раза большие размеры. Причиной тому была экспрессия гена гормона роста, в результате которой у мыши синтезировался соответствующий гормон, который и определил ее гигантские размеры.
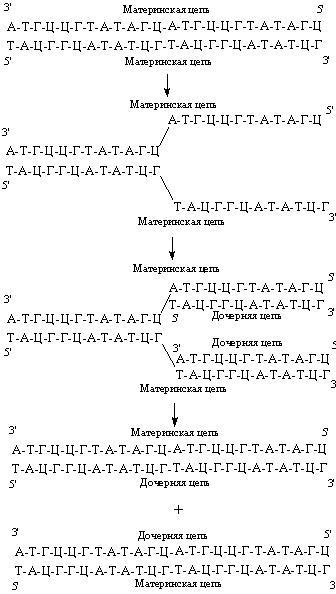
Рис. 44. В результате репликации образуются две дочерние молекулы ДНК.
Репликация ДНК
В основе репликации, или удвоения, ДНК лежит принцип комплементарности. Репликация начинается с разделения цепей, каждая из которых становится матрицей, определяющей нуклеотидную последовательность новой комплементарной цепи (рис. 44). В результате репликации образуются две дочерние молекулы ДНК, нуклеотидные последовательности которых идентичны между собой и с материнской молекулой ДНК. При этом две дочерние молекулы ДНК содержат одну вновь синтезированную (дочернюю) и одну материнскую цепи ДНК.
Субстратом для синтеза ДНК являются дезоксинуклеотид-5’-трифосфаты: дАТФ, дГТФ, дЦТФ и дТТФ. Уравнение этой реакции в простейшей виде выглядит так:
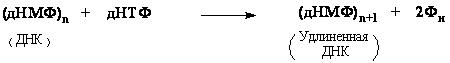
Данную реакцию катализирует фермент ДНК-полимераза. Она последовательно добавляет нуклеотиды к 3’-концу полинуклеотидной цепи. Следовательно, синтез новой цепи происходит в направлении от 5’-конца к 3’-концу. ДНК-полимераза может только наращивать цепь ДНК, начать же синтез ДНК с нуля она не может, т.е. для ее работы требуется цепь-затравка. ДНК-полимераза способна удлинять цепь только в присутствии цепи, играющей роль матрицы (рис. 45.). Нуклеотиды присоединяются к цепи-затравке в соответствии с принципом комплементарности, напротив аденина всегда будет встроен тимин, а напротив гуанина – цитозин.
Итогом репликации является образование двух дочерних молекул ДНК, являющихся точными копиями материнской. Благодаря этому возможна передача идентичной генетической информации от клетки к клетке.
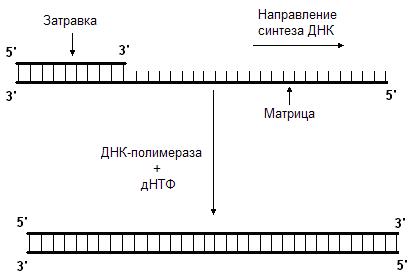
Рис. 45. ДНК-полимераза наращивает цепь в направлении 5’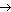 3’.
3’.
Для этого ей нужны матрица и затравка
Транскрипция
В результате транскрипции происходит синтез цепи РНК, нуклеотидная последовательность которой комплементарна одной из цепей ДНК; при этом генетическая информация, содержащаяся в ДНК, передается молекуле РНК. В результате транскрипции образуются три класса РНК: иРНК, тРНК, рРНК. Поскольку в основе транскрипции лежит принцип комплементарности, этот процесс имеет определенное сходство с репликацией. Однако между ними существует и важное различие. Если в процессе репликации копируется вся молекула ДНК, то при транскрипции транскрибируется только ее незначительная часть. Катализирует синтез РНК фермент РНК-полимераза. Транскрибируемый участок ДНК ограничен со стороны 3’-конца промотором, – участком с которым связывается РНК-полимераза, со стороны 5’-конца – терминатором – участком, в котором прерывается синтез РНК. Последовательность ДНК, ограниченная промотором и терминатором, представляет собой единицу транскрипции – транскриптон (рис. 46).
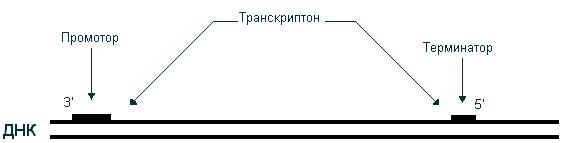
Рис. 46. Транскриптон
РНК-полимераза в качестве субстратов для синтеза РНК использует рибонуклеозид-5’-фосфаты (АТФ, ГТФ, ЦТФ и УТФ). Она удлиняет цепь, присоединяя нуклеотиды к 3’-концу, т.е. так же, как и ДНК-полимераза, наращивает цепь в направлении5’®3’. Этот фермент требует для своей работы в качестве матрицы одну из цепей ДНК. Из двух цепей ДНК транскрибируется только одна. В отличие от ДНК-полимеразы, РНК-полимераза не нуждается в затравке. Удлинение цепи РНК описывается уравнением:

Нуклеотиды присоединяются к цепи в соответствии с принципом комплементарности. Напротив аденина матричной цепи ДНК в растущую цепь РНК всегда будет встроен урацил, напротив гуанина – цитозин, напротив тимина – аденин и напротив цитозина – гуанин соответственно. Т.о., образовавшаяся в результате транскрипции РНК комплементарна матричной цепи ДНК (рис. 47).
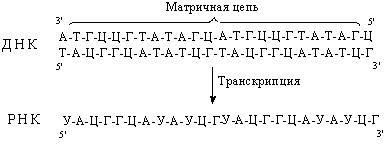
Рис. 47. Образовавшаяся в результате транскрипции РНК комплементарна матричной цепи ДНК
В процессе транскрипции выделяют три стадии: инициацию, элонгацию и терминацию. На стадии инициации РНК-полимераза, взаимодействуя с промотором, вызывает расхождение цепей ДНК и начинает синтез молекулы РНК. В ходе элонгации новосинтезированная цепь РНК образует (за счет спаривания ее оснований с основаниями матричной цепи ДНК) короткие отрезки гибридной двойной спирали ДНК – РНК, которые необходимы для правильного считывания цепи ДНК. Как только РНК-полимераза достигнет терминирующих последовательностей, запускается последняя стадия – терминация. По ее завершении происходит освобождение вновь синтезированной цепи РНК (рис. 48).
Рис. 48. Стадии транскрипции
Синтезированные молекулы РНК могут подвергаться посттранскрипционным ковалентным модификациям, так называемому процессингу (созреванию). Наиболее ярко процессинг выражен у эукариот.
Процессинг РНК
Рассмотрим процессинг иРНК эукариот.
иРНК синтезируется в виде предшественника пре-иРНК. Модификация пре-иРНК начинается на стадии элонгации, в это время происходит кэпирование 5’-конца. Остаток ГТФ присоединяется своим 5’-концом к 5’-концу пре-иРНК с образованием так называемого кэпа. После завершения транскрипции к 3’-концу присоединяется полиА-последовательность, состоящая из 100 – 200 последовательно соединенных адениновых нуклеотидов. В составе пре-иРНК могут содержаться интроны и экзоны. Интроны при созревании РНК вырезаются, концы экзонов соединяются друг с другом. Этот процесс называется сплайсингом. В результате сплайсинга зрелая молекула иРНК становится приблизительно в 4 раза короче пре-иРНК. Этапы процессинга иРНК представлены на рис. 49. Процессинг для иРНК прокариот не характерен.
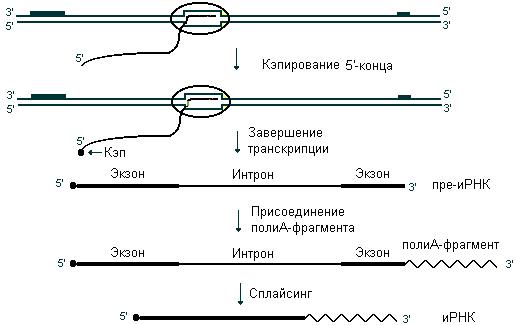
Рис. 49. Процессинг иРНК
рРНК и тРНК также синтезируются в виде более длинных предшественников, которые затем расщепляются и модифицируются. У прокариот рРНК представлены тремя молекулами: 16S-рРНК, 23S-рРНК и 5S-рРНК, у эукариот – четырьмя: 18S-рРНК, 28S-рРНК, 5,8S-рРНК и 5S-рРНК. рРНК как прокариот, так и эукариот образуются из предшественников – пре-рРНК. Предшественник расщепляется, образуя индивидуальные рРНК (рис. 50). рРНК и тРНК не содержат кэпа и полиА-последовательности. У эукариот рРНК и тРНК могут подвергаться сплайсингу. В процессе созревания у некоторых предшественников тРНК наряду с удалением концевых последовательностей может происходить и присоединение нуклеотидных последовательностей, играющих важную роль в их функционировании.
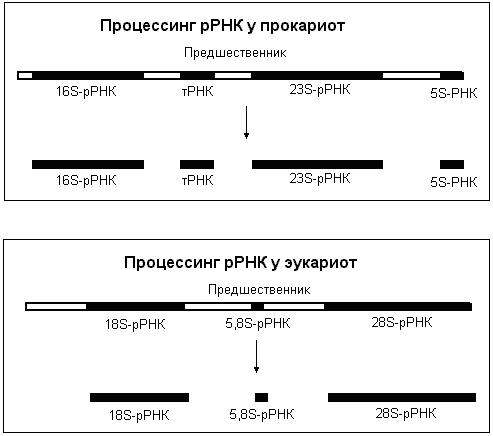
Рис.50. Процессинг рРНК.
Обратная транскрипция
Синтез ДНК при использовании в качестве матрицы РНК носит название обратной транскрипции. Данный процесс катализирует фермент обратная транскриптаза или ревертаза. Существование обратных транскриптаз в составе РНК-содержащих вирусов было показано Г.Темином и Д. Балтимором. Обнаружение обратной транскриптазы позволило ответить на вопрос: как генетическая информация РНК-содержащих вирусов может включиться в ДНК клетки-хозяина. Процесс обратной транскрипции, катализируемый ревертазой, и последующая интеграция генетического материала в геном клетки хозяина представлены на рис. 51. В процессе обратной транскрипции вначале образуется дуплекс РНК – ДНК, затем РНК в составе этого дуплекса разрушается, синтезированная цепь ДНК далее служит матрицей для синтеза второй цепи ДНК. На заключительной стадии происходит интеграция ДНК, синтезированной в результате обратной транскрипции, в ДНК клетки-хозяина. Обратные транскриптазы способны синтезировать ДНК, комплементарную самым различным РНК. Благодаря этой особенности ревертаза нашла широкое применение в научных исследованиях. С помощью обратной транскриптазы можно получить, например, искусственный ген, используя в качестве матрицы иРНК.
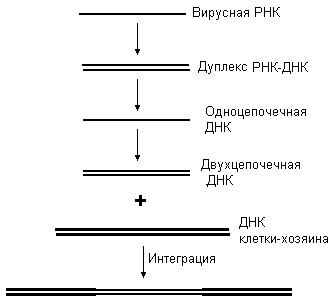
Рис. 51. Обратная транскрипция