§ 12. КИНЕТИКА ФЕРМЕНТАТИВНЫХ РЕАКЦИЙ
Кинетика ферментативных реакций – наука о скоростях ферментативных реакций, их зависимости от различных факторов. Скорость ферментативной реакции определяется химическим количеством прореагировавшего субстрата или образовавшегося продукта реакции в единицу времени в единице объема при определенных условиях:
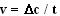 ,
,
где v – скорость ферментативной реакции, 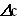 – изменение концентрации субстрата или продукта реакции, t – время.
– изменение концентрации субстрата или продукта реакции, t – время.
Скорость ферментативной реакции зависит от природы фермента, которая определяет его активность. Чем выше активность фермента, тем выше скорость реакции. Активность фермента определяют по скорости реакции, катализируемой ферментом. Мерой активности фермента является одна стандартная единица активности фермента. Одна стандартная единица активности фермента – это такое количество фермента, которое катализирует превращение 1 мкмоль субстрата за 1 минуту.
В процессе ферментативной реакции фермент (Е) взаимодействует с субстратом (S), в результате образуется фермент-субстратный комплекс, который затем распадается с высвобождением фермента и продукта (Р) реакции:
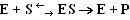
Скорость ферментативной реакции зависит от многих факторов: от концентрации субстрата и фермента, температуры, рН среды, наличия различных регуляторных веществ, способных увеличивать или снижать активность ферментов.
Интересно знать! Ферменты используются в медицине для диагностики различных заболеваний. При инфаркте миокарда вследствие повреждения и распада сердечной мышцы в крови резко возрастает содержание ферментов аспартаттрансаминазы и аланинаминотрансферазы. Выявление их активности позволяет диагностировать данное заболевание.
Влияние концентрации субстрата и фермента на скорость ферментативной реакции
Рассмотрим влияние концентрации субстрата на скорость ферментативной реакции (рис. 30.). При низких концентрациях субстрата скорость прямо пропорциональна его концентрации, далее с ростом концентрации скорость реакции увеличивается медленнее, а при очень высоких концентрациях субстрата скорость практически не зависит от его концентрации и достигает своего максимального значения (Vmax). При таких концентрациях субстрата все молекулы фермента находятся в составе фермент-субстратного комплекса, и достигается полное насыщение активных центров фермента, именно поэтому скорость реакции в этом случае практически не зависит от концентрации субстрата.
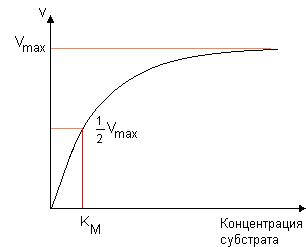
Рис. 30. Зависимость скорости ферментативной реакции от концентрации субстрата
График зависимости активности фермента от концентрации субстрата описывается уравнением Михаэлиса – Ментен, которое получило свое название в честь выдающихся ученых Л.Михаэлиса и М.Ментен, внесших большой вклад в исследование кинетики ферментативных реакций,
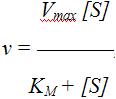
где v – скорость ферментативной реакции; [S] – концентрация субстрата; KM – константа Михаэлиса.
Рассмотрим физический смысл константы Михаэлиса. При условии, что v = ½ Vmax, получаем KM = [S]. Таким образом, константа Михаэлиса равна концентрации субстрата, при которой скорость реакции равна половине максимальной.
Скорость ферментативной реакции зависит и от концентрации фермента (рис. 31). Эта зависимость носит прямолинейный характер.
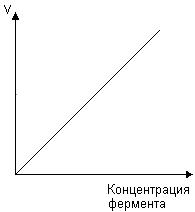
Рис. 31. Зависимость скорости ферментативной реакции от концентрации фермента
Влияние температуры на скорость ферментативной реакции
Зависимость скорости ферментативной реакции от температуры представлена на рис. 32.
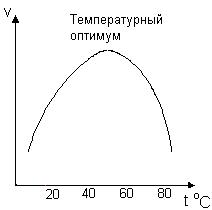
Рис. 32. Зависимость скорости ферментативной реакции от температуры.
При низких температурах (приблизительно до 40 – 50 оС) повышение температуры на каждые 10 оС в соответствии с правилом Вант-Гоффа сопровождается увеличением скорости химической реакции в 2 – 4 раза. При высоких температурах более 55 – 60 оС активность фермента резко снижается из-за его тепловой денатурации, и, как следствие этого, наблюдается резкое снижение скорости ферментативной реакции. Максимальная активность ферментов наблюдается обычно в пределах 40 – 60 оС. Температура, при которой активность фермента максимальна, называется температурным оптимумом. Температурный оптимум ферментов термофильных микроорганизмов находится в области более высоких температур.
Влияние рН на скорость ферментативной реакции
График зависимости ферментативной активности от рН представлен на рис. 33.
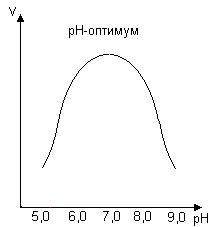
Рис. 33. Влияние рН на скорость ферментативной реакции
График зависимости от рН имеет колоколообразную форму. Значение рН, при котором активность фермента максимальна, называется рН-оптимумом фермента. Значения рН-оптимума для различных ферментов колеблются в широких пределах.
|
Фермент |
рН-оптимум |
|
Пепсин |
1,5 |
|
Фосфатаза |
5,8 |
|
Уреаза |
6,7 |
|
Трипсин |
7,7 |
|
Каталаза |
7,6 |
|
Аргиназа |
9,7 |
Характер зависимости ферментативной реакции от рН определяется тем, что этот показатель оказывает влияние на:
a) ионизацию аминокислотных остатков, участвующих в катализе,
b) ионизацию субстрата,
c) конформацию фермента и его активного центра.
Ингибирование ферментов
Скорость ферментативной реакции может быть снижена действием ряда химических веществ, называемых ингибиторами. Некоторые ингибиторы являются для человека ядами, например, цианиды, другие – используются в качестве лекарственных препаратов.
Ингибиторы можно разделить на два основных типа: необратимые и обратимые. Необратимые ингибиторы (I) связываются с ферментом с образованием комплекса, диссоциация которого с восстановлением активности фермента невозможна:
E + I  EI.
EI.
Примером необратимого ингибитора является диизопропилфторфосфат (ДФФ). ДФФ ингибирует фермент ацетилхолинэстеразу, играющего важную роль в передаче нервного импульса. Этот ингибитор взаимодействует с серином активного центра фермента, блокируя тем самым активность последнего. Вследствие этого нарушается способность отростков нервных клеток нейронов проводить нервный импульс. ДФФ является одним из первых веществ нервно-паралитического действия. На его основе создан ряд относительно нетоксичных для человека и животных инсектицидов - веществ, ядовитых для насекомых.
Обратимые ингибиторы, в отличие от необратимых, при определенных условиях могут быть легко отделены от фермента. Активность последнего при этом восстанавливается:
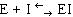 .
.
Среди обратимых ингибиторов выделяют конкурентные и неконкурентные ингибиторы.
Конкурентный ингибитор, являясь структурным аналогом субстрата, взаимодействует с активным центром фермента и таким образом перекрывает доступ субстрата к ферменту. При этом ингибитор не подвергается химическим превращениям и связывается с ферментом обратимо. После диссоциации комплекса EI фермент может связаться либо с субстратом и преобразовать его, либо с ингибитором (рис. 34.). Поскольку и субстрат и ингибитор конкурируют за место в активном центре, такое ингибирование называется конкурентным.
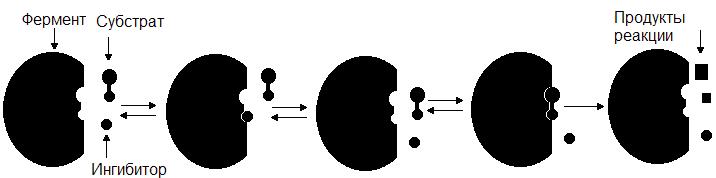
Рис. 34. Механизм действия конкурентного ингибитора.
Конкурентные ингибиторы используются в медицине. Для борьбы с инфекционными болезнями ранее широко применялись сульфаниламидные препараты. Они близки по своей структуре к пара-аминобензойной кислоте (ПАБК), необходимому фактору роста многих патогенных бактерий. ПАБК является предшественником фолиевой кислоты, которая служит кофактором ряда ферментов. Сульфаниламидные препараты выступают в качестве конкурентного ингибитора ферментов синтеза фолиевой кислоты из ПАБК и тем самым подавляют рост и размножение патогенных бактерий.
Неконкурентные ингибиторы по структуре не сходны с субстратом и при образовании EI взаимодействуют не с активным центром, а с другим участком фермента. Взаимодействие ингибитора с ферментом приводит к изменению структуры последнего. Образование EI-комплекса является обратимым, поэтому после его распада фермент вновь способен атаковать субстрат (рис. 35).
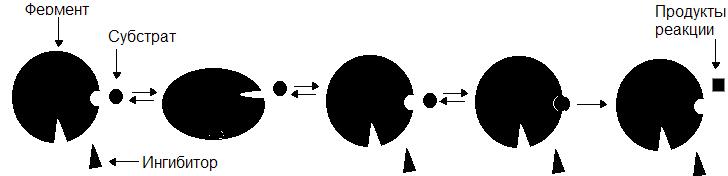
Рис. 35. Механизм действия неконкурентного ингибитора
В качестве неконкурентного ингибитора может выступать цианид CN-. Он связывается с ионами металлов, входящими в состав простетических групп и подавляет активность этих ферментов. Отравления цианидами крайне опасны. Они могут привести к летальному исходу.
Аллостерические ферменты
Термин «аллостерический» происходит от греческих слов allo – другой, stereo – участок. Таким образом, аллостерические ферменты наряду с активным центром имеют другой центр, называемый аллостерический центр (рис. 36). С аллостерическим центром связываются вещества, способные изменять активность ферментов, эти вещества называют аллостерическими эффекторами. Эффекторы бывают положительными – активирующими фермент, и отрицательными – ингибирующими, т.е. снижающими активность фермента. Некоторые аллостерические ферменты могут подвергаться действию двух и более эффекторов.
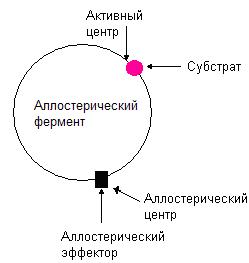
Рис. 36. Структура аллостерического фермента.
Регуляция мультиферментных систем
Некоторые ферменты действуют согласованно, объединяясь в мультиферментные системы, в которых каждый фермент катализирует определенную стадию метаболитического пути:
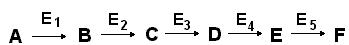
В мультиферментной системе есть фермент, который определяет скорость всей последовательности реакций. Этот фермент, как правило, бывает аллостерическим и находится в начале матаболитического пути. Он способен, получая различные сигналы, как повышать, так и понижать скорость катализируемой реакции, тем самым регулируя скорость всего процесса.