§ 8. ПРОСТРАНСТВЕННАЯ ОРГАНИЗАЦИЯ БЕЛКОВОЙ МОЛЕКУЛЫ
Первичная структура
Под первичной структурой белка понимают количество и порядок чередования аминокислотных остатков, соединенных друг с другом пептидными связями, в полипептидной цепи.
Полипептидная цепь на одном конце содержит свободную, не участвующую в образовании пептидной связи, NH2-группу, этот участок обозначается как N–конец. На противоположной стороне располагается свободная, не участвующая в образовании пептидной связи, НООС-группа, это – С-конец. За начало цепи принимается N-конец, именно с него начинается нумерация аминокислотных остатков:
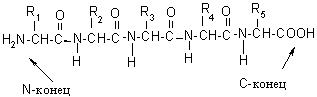
Аминокислотную последовательность инсулина установил Ф. Сэнгер (Кембриджский университет). Этот белок состоит из двух полипептидных цепей. Одна цепь состоит из 21 аминокислотного остатка, другая цепь – из 30. Цепи связаны двумя дисульфидными мостиками (рис.6).

Рис. 6. Первичная структура инсулина человека
На расшифровку этой структуры было затрачено 10 лет (1944 – 1954 гг.). В настоящее время первичная структура определена у многих белков, процесс ее определения автоматизирован и не представляет собой серьезную проблему для исследователей.
Информация о первичной структуре каждого белка закодирована в гене (участке молекулы ДНК) и реализуется в ходе транскрипции (переписывании информации на мРНК) и трансляции (синтеза полипептидной цепи). В связи с этим можно установить первичную структуру белка также по известной структуре соответствующего гена.
По первичной структуре гомологичных белков можно судить о таксономическом родстве видов. К гомологичным белкам относятся те белки, которые у разных видов выполняют одинаковые функции. Такие белки имеют сходные аминокислотные последовательности. Например, белок цитохром С у большинства видов имеет относительную молекулярную массу около 12500 и содержит около 100 аминокислотных остатков. Различия в первичной структуре цитохрома С двух видов пропорциональны филогенетическому различию между данными видами. Так цитохромы С лошади и дрожжей отличаются по 48 аминокислотным остаткам, курицы и утки – по двум, цитохромы же курицы и индейки идентичны.
Вторичная структура
Вторичная структура белка формируется вследствие образования водородных связей между пептидными группами. Различают два типа вторичной структуры: α-спираль и β-структура (или складчатый слой). В белках могут присутствовать также участки полипептидной цепи, не образующие вторичную структуру.
α-Спираль по форме напоминает пружину. При формировании α-спирали атом кислорода каждой пептидной группы образует водородную связь с атомом водорода четвертой по ходу цепи NH-группы:

Каждый виток спирали связан со следующим витком спирали несколькими водородными связями, что придает структуре значительную прочность. α-Спираль обладает следующими характеристиками: диаметр спирали 0,5 нм, шаг спирали – 0,54 нм, на один виток спирали приходится 3,6 аминокислотных остатка (рис. 7).

Рис. 7. Модель a-спирали, отражающая ее количественные характеристики
Боковые радикалы аминокислот направлены наружу от 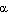 -спирали (рис. 8).
-спирали (рис. 8).
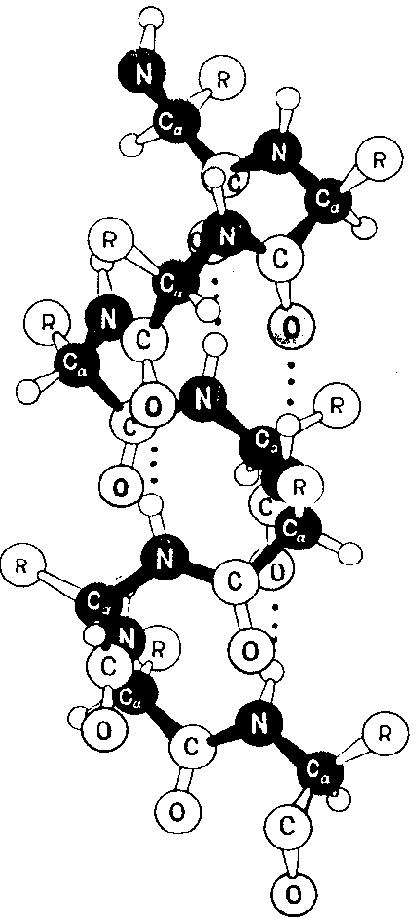
Рис. 8. Модель -спирали, отражающая пространственное расположение боковых радикалов
Из природных L-аминокислот может быть построена как правая, так и левая -спираль. Для большинства природных белков характерна правая спираль. Из D-аминокислот также можно построить как левую, так и правую спираль. Полипептидная же цепь, состоящая из смеси D-и L-аминокислотных остатков, не способна образовывать спираль.
Некоторые аминокислотные остатки препятствуют образованию α-спирали. Например, если в цепи подряд расположено несколько положительно или отрицательно заряженных аминокислотных остатков, такой участок не примет α-спиральной структуры из-за взаимного отталкивания одноименно заряженных радикалов. Затрудняют образование -спирали радикалы аминокислотных остатков, имеющих большие размеры. Препятствием для образования α-спирали, является также наличие в полипептидной цепи остатков пролина (рис. 9). В остатке пролина при атоме азота, образующем пептидную связь с другой аминокислотой, нет атома водорода.

Рис. 9. Остаток пролина препятствует образованию -спирали
Поэтому остаток пролина, входящий в состав полипептидной цепи, не способен образовывать внутрицепочечную водородную связь. Кроме того, атом азота в пролине входит в состав жесткого кольца, что делает невозможным вращение вокруг связи N – C и образование спирали.
Кроме α-спирали описаны и другие типы спиралей. Однако они встречаются редко, в основном на коротких участках.
Образование водородных связей между пептидными группами соседних полипептидных фрагментов цепей приводит к формированию β-структуры, или складчатого слоя:

В отличие от α-спирали складчатый слой имеет зигзагообразную форму, похожую на гармошку (рис. 10).
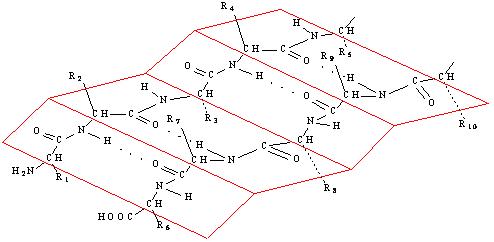
Рис. 10. β-Структура белка
Различают параллельные и антипараллельные складчатые слои. Параллельные β-структуры образуются между участками полипептидной цепи, направления которых совпадают:

Антипаралельные β-структуры образуются между противоположно направленными участками полипептидной цепи:

β-Структуры могут формироваться более чем между двумя полипептидными цепями:

В составе одних белков вторичная структура может быть представлена только α-спиралью, в других – только β-структурами (параллельными, или антипараллельными, или и теми, и другими), в третьих наряду с α-спирализованными участками могут присутствовать и β-структуры.
Третичная структура
У многих белков вторичноорганизованные структуры (α-спирали,  -структуры) свернуты определенным образом в компактную глобулу. Пространственная организация глобулярных белков носит название третичной структуры. Таким образом, третичная структура характеризует трехмерное расположение участков полипептидной цепи в пространстве. В формировании третичной структуры принимают участие ионные и водородные связи, гидрофобные взаимодействия, ван-дер-ваальсовы силы. Стабилизируют третичную структуру дисульфидные мостики.
-структуры) свернуты определенным образом в компактную глобулу. Пространственная организация глобулярных белков носит название третичной структуры. Таким образом, третичная структура характеризует трехмерное расположение участков полипептидной цепи в пространстве. В формировании третичной структуры принимают участие ионные и водородные связи, гидрофобные взаимодействия, ван-дер-ваальсовы силы. Стабилизируют третичную структуру дисульфидные мостики.
Третичная структура белков определяется их аминокислотной последовательностью. При ее формировании связи могут возникать между аминокислотами, расположенными в полипептидной цепи на значительном расстоянии. У растворимых белков полярные радикалы аминокислот, как правило, оказываются на поверхности белковых молекул и реже – внутри молекулы, гидрофобные радикалы оказываются компактно упакованными внутри глобулы, образуя гидрофобные области.
В настоящее время третичная структура многих белков установлена. Рассмотрим два примера.
Миоглобин
Миоглобин – кислород-связывающий белок с относительной массой 16700. Его функция – запасание кислорода в мышцах. В его молекуле имеется одна полипептидная цепь, состоящая из 153 аминокислотных остатков, и гемогруппа, играющая важную роль в связывании кислорода.
Пространственная организация миоглобина установлена благодаря работам Джона Кендрью и его коллег (рис. 11). В молекуле этого белка присутствуют 8 α-спиральных участков, на их долю приходится 80 % всех аминокислотных остатков. Молекула миоглобина очень компактна, внутри нее может уместиться всего четыре молекулы воды, почти все полярные радикалы аминокислот расположены на внешней поверхности молекулы, большая часть гидрофобных радикалов расположена внутри молекулы, вблизи поверхности находится гем – небелковая группа, ответственная за связывание кислорода.
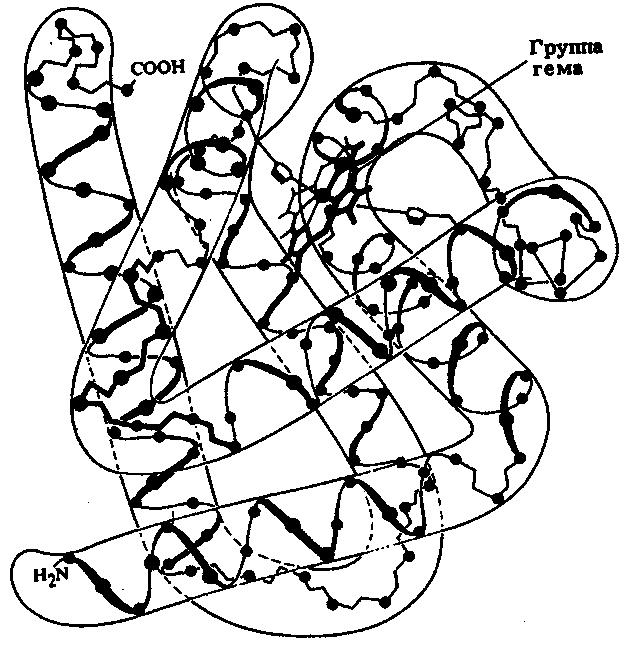
Рис.11. Третичная структура миоглобина
Рибонуклеаза
Рибонуклеаза – глобулярный белок. Она секретируется клетками поджелудочной железы, это – фермент, катализирующий расщепление РНК. В отличие от миоглобина, в молекуле рибонуклеазы имеется очень мало α-спиральных участков и достаточно большое число сегментов, находящихся в β-конформации. Прочность третичной структуре белка придают 4 дисульфидные связи.
Четвертичная структура
Многие белки состоят из нескольких, двух или более, белковых субъединиц, или молекул, обладающих определенной вторичной и третичной структурами, удерживаемых вместе при помощи водородных и ионных связей, гидрофобных взаимодействий, ван-дер-ваальсовых сил. Такая организация белковых молекул носит название четвертичной структуры, а сами белки называют олигомерными. Отдельная субъединица, или белковая молекула, в составе олигомерного белка называется протомером.
Число протомеров в олигомерных белках может варьировать в широких пределах. Например, креатинкиназа состоит из 2 протомеров, гемоглобин – из 4 протомеров, РНК-полимераза E.coli – фермент, ответственный за синтез РНК, – из 5 протомеров, пируватдегидрогеназный комплекс – из 72 протомеров. Если белок состоит из двух протомеров, его называют димером, четырех – тетрамером, шести – гексамером (рис. 12). Чаще в молекуле олигомерного белка содержится 2 или 4 протомера. В состав олигомерного белка могут входить одинаковые или различные протомеры. Если в состав белка входят два идентичных протомера, то это – гомодимер, если разные – гетеродимер.

Рис. 12. Олигомерные белки
Рассмотрим организацию молекулы гемоглобина. Основная функция гемоглобина заключается в транспорте кислорода из легких в ткани и углекислого газа в обратном направлении. Его молекула (рис. 13) состоит из четырех полипептидных цепей двух различных типов – двух α-цепей и двух β-цепей и гема. Гемоглобин является белком, родственным миоглобину. Вторичная и третичная структуры миоглобина и протомеров гемоглобина очень сходны. Каждый протомер гемоглобина содержит, как и миоглобин, 8 α-спирализованных участков полипептидной цепи. При этом надо отметить, что в первичных структурах миоглобина и протомера гемоглобина идентичны только 24 аминокислотных остатка. Следовательно, белки, значительно отличающиеся по первичной структуре, могут иметь сходную пространственную организацию и выполнять сходные функции.
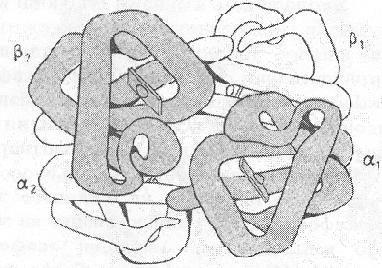
Рис. 13. Структура гемоглобина