РАЗДЕЛ 4. ФИЗИОЛОГИЯ ДЫХАНИЯ
Дыхание как физиологическая функция, включает:
1. Газообмен – совокупность процессов, обеспечивающих поступление О2 в организм, использование его в окислительно-восстановительных процессах и удаление продуктов окисления (через устьица, клеточные стенки, покровные ткани).
2. Темновое дыхание – совокупность окислительно-восстановительных процессов разного типа и назначения, в ходе которых сложные органические вещества распадаются на более простые, а высвобождаемая энергия в сопряженных реакциях аккумулируется в лабильных фосфорных связях (АТФ) и используется во всех эндергонических процессах клетки и растительного организма в целом. Различают следующие виды клеточного дыхания:
а) сопряженное с запасанием энергии в утилизируемой форме;
б) свободное окисление – перенос электронов, присоединение О2, не связанные с энергетической функцией (терморегуляция, конструктивная и деструктивная функции, детоксикация ксенобиотиков, косвенное запасание Е).
3. фотодыхание(выделение СО2 на свету в процессах, связанных с фотосинтезом).
Главными функциямидыхания являются:
а) мобилизация энергетических ресурсов дыхательного субстрата путем трансформации стабильных соединений в активную форму (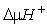 , АТФ), доступную для утилизации, то есть использование в различных видах биологически полезной работы;
, АТФ), доступную для утилизации, то есть использование в различных видах биологически полезной работы;
б) мобилизация пластических резервов дыхательного субстрата, путем образования многочисленных промежуточных продуктов, то есть высвобождение («добыча») активных строительных материалов (блоков);
в) терморегуляционная (теплопродукция) - рассеяние энергии в виде тепла;
г) защитная – расщепление (детоксикация) вредных веществ, то есть защита от неблагоприятных факторов.
Компонентами дыхания в обеспечении энергетической и пластической функций являются:
1) дыхание роста (ДР) – затраты на процессы роста и развитие организма;
2) дыхание поддержания (ДП):
а) ресинтез веществ, претерпевающих обновление в процессе метаболизма (ферментные белки, РНК, липиды мембран и др.);
б) поддержание в клетке необходимой концентрации ионов, рН;
г) поддержание уровня метаболитов против градиента концентрации;
д) поддержание функционально-активного состояния структур.
Математически описывается формулой:
Д = хДр + уДп,
где x и y – коэффициенты затрат.
Процесс дыхания протекает в 4 этапа:
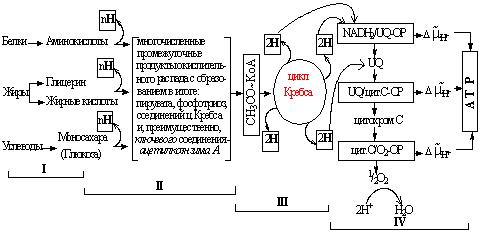
I. Подготовительный – деполимеризация макромолекул.
II. Частичное анаэробное окисление и унификация дыхательного субстрата. Часть энергии мобилизуется в форме восстановленных коферментов специфических ферментов дыхания – дегидрогеназ, а также в форме АТР в 2 реакциях субстратного фосфорилирования гликолиза. В ходе окисления образуются многочисленные промежуточные продукты – предшественники реакций специфического синтеза. Для дальнейшего окисления дыхательный субстрат унифицируется, то есть образуется сравнительно небольшое количество соединений, главным образом ацетилкоэнзим А – ключевое соединение реакций синтеза и распада. Эти соединения могут непосредственно включаться в цикл Кребса, который, таким образом, является универсальным путем окисления всех видов субстрата.
III. Аэробное окисление дыхательного субстрата в цикле Кребса. Энергия мобилизуется в 4 окислительных реакциях в форме восстановленных дегидрогеназ: изоцитрат-, оксоглутарат-, сукцинат- и малат-, а также в реакции субстратного фосфорилирования (в ходе окислительного декарбоксилирования оксоглутаровой кислоты до сукцината). Промежуточные продукты, в основном кетокислоты, используются в реакциях специфического синтеза.
IV. Окисление электронов, поставляемых дегидрогеназами в дыхательной цепи митохондрий. Электроны, «мобилизованные» специфическими дегидрогеназами на предыдущих этапах, могут передаваться на дыхательную цепь, где происходит их энергетическая разгрузка с трансформацией энергии в форму протонного градиента внутренних мембран митохондрий. Дыхательная цепь состоит из трех основных «энергопреобразующих блоков» – комплексов ферментов: никотинамидадениндинуклеотид/убихинон-оксидоредуктаза (NADH/UQ-ОР), убихинон/цитохром «с»-оксидоредуктаза ((UQ/цит. с-ОР) и цитохром «с»/кислород-оксидоредуктаза. В зависимости от энергетического потенциала, передаваемого на цепь электрона, он может быть передан на уровне 1-го, 2-го или 3-го комплекса. В связи с этим может запастись большее или меньшее количество энергии в форме . Протонный градиент как мембранная форма энергетической валюты может расходоваться в сопряженных реакциях на синтез АТФ с участием АТР-синтазного комплекса F0F1-типа.